

























Adults and larvae of many species in this family appear to be facultative predators of other small arthropods, in that they feed on both floral tissues as well as on thrips and mites that live in flowers. However, some species are almost certainly solely phytophagous, a few being univoltine in flowers of particular plant species (Tyagi, et al., 2008), whereas in the warmer parts of the world, a considerable number of species are obligate predators (Hoddle, 2003).
In Australia, the main aeolothripid genus, Desmothrips, includes many species that are common in a wide range of flowers, and most of these seem to be facultative predators. This also appears to be true of the many species in the northern hemisphere genus Aeolothrips. However, in both of these genera some species appear to be predatory, and a few of them are distinctive ant-mimics. These include Aeolothrips albicinctus in Europe, Aeolothrips bicintus and related species in North America, as well as Desmothrips reedi in southern Australia. Ant-mimicry has also arisen amongst other members of this family, including Franklinothrips orizabensis has been employed as a biocontrol agent against thrips pests in European greenhouses. Species of the tropical genus Mymarothrips are also obligate predators, but have a body form and wings that ressemble those of small chalcid wasps. In contrast, Cycadothrips species are phytophagous, breeding only in the males cones of Macrozamia cycads in Australia (Mound & Terry, 2001), and Dactuliothrips species also seem to be phytophagous breeding in California particularly in the flowers of Yucca species (Hoddle et al., 2012).
It is probable that in all species of Aeolothripidae the second instar larvae produce a silken cocoon within which to pupate (Hoddle et al., 2001), and such cocoons usually occur at ground level.
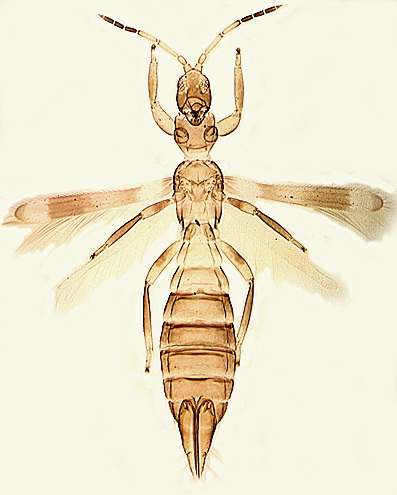
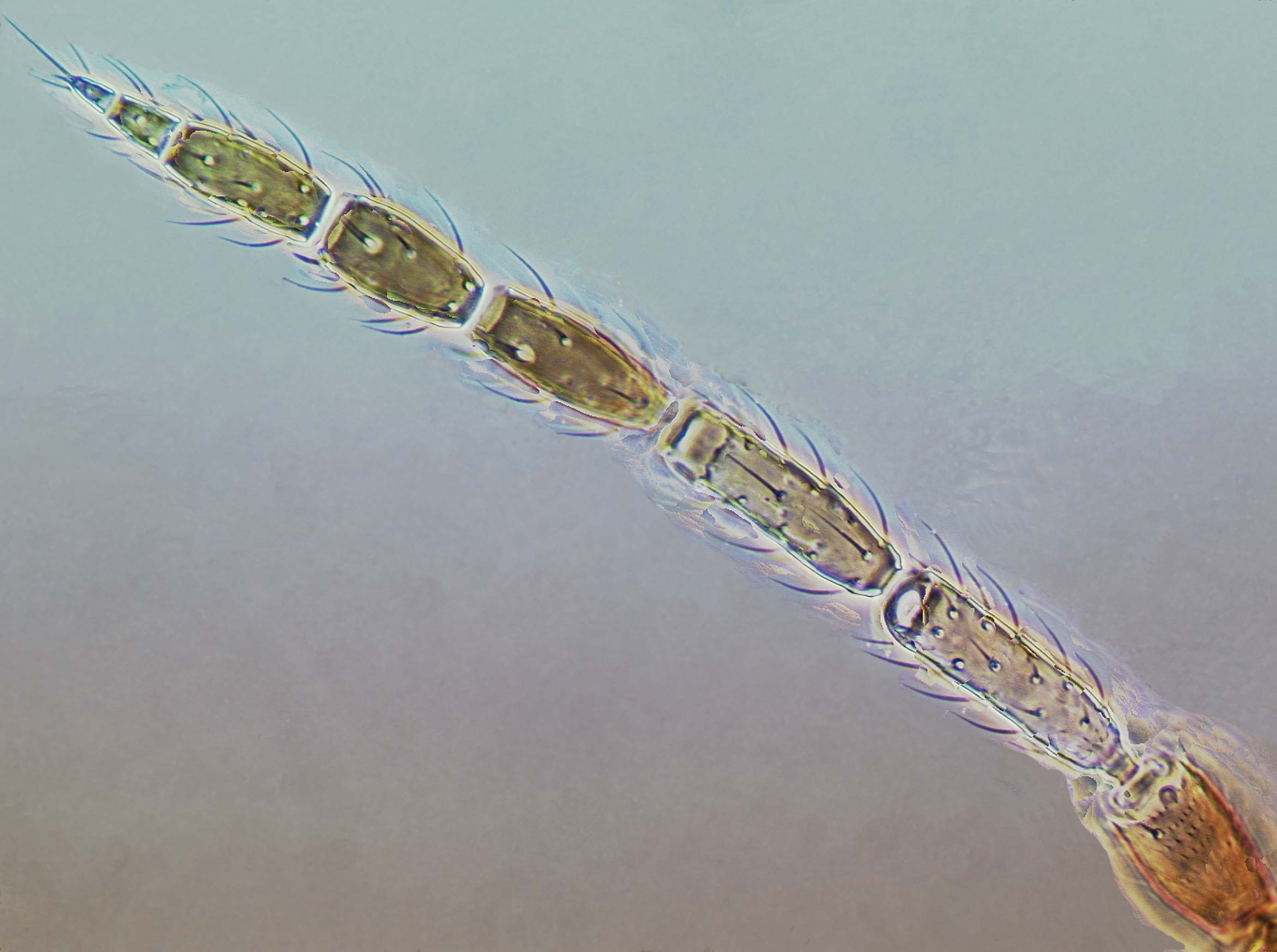
Species of Aeolothripidae all have nine antennal segments, of which at least segments VII – IX are broadly connate and form a single unit. The sensoria on segments III and IV are generally linear and longitudinal in the apical portion of these segments but often curling around the segmental apex. Exceptions to this occur in Rhipidothrips, in which the sensoria are interpreted as being reduced to the apical curve except in R. gratiosus. Also in Orothrips, Dactuliothrips, and Cycadothrips, these two segments each bear two sensoria.
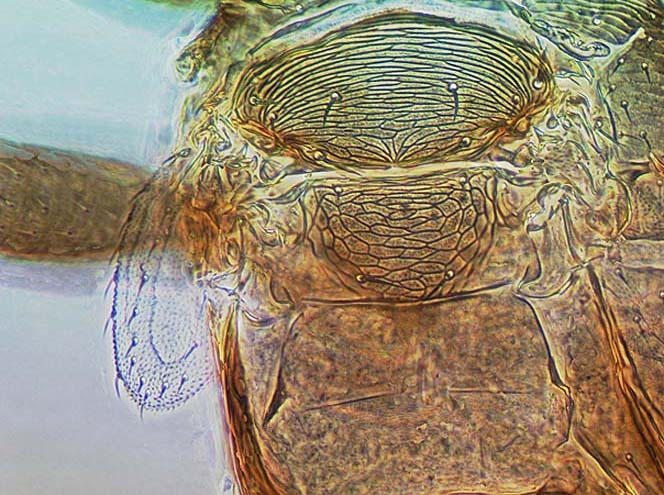
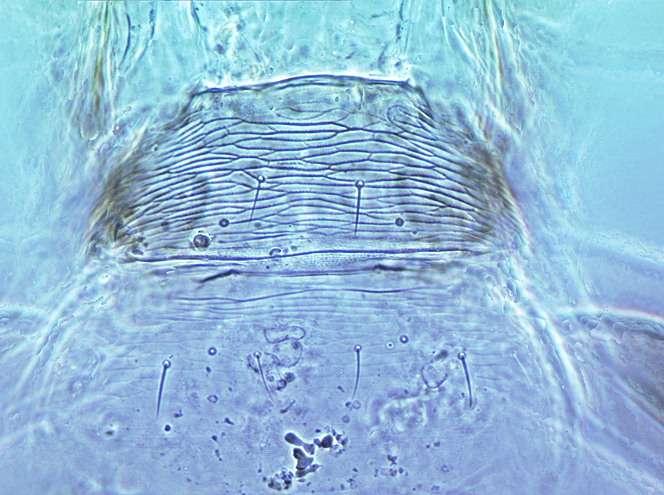
Aeolothripidae are distinguished from the Melanthripidae by the complete loss of sternite VIII. The pair of lobes that represent this sternite in Melanthripidae each bear two setae, and these two pairs of setae are retained submarginally on sternite VII of Aeolothripidae. In the head, the tentorial bridge is well-developed, but the anterior tentorial arms are weaker than in Melanthripidae. The forewings often have transverse or longitudinal dark bands, and are broadly rounded at the apex, with several cross-veins. Most Aeolothripidae have only short setae on the head and pronotum, but Dactuliothrips species have many prominent setae as in many Melanthripidae.
Worldwide, almost 200 species of Aeolothripidae are recognised, in 23 genera, although the distribution of species within genera is very asymmetric. The genus Aeolothrips includes just over 90 species, eight genera each include a single species, a further seven each include less than five species, and only three genera include between 10 and 15 species. For details see Mound’s Thysanoptera pages.
The genus Aeolothrips is sometimes restricted (Bhatti, 2006) to a single ground-living species, A. albicinctus, in which the first abdominal tergite is narrowed and bears numerous transverse striae; the other 89 species being treated in Coleothrips. However, a similar transversely striate first abdominal tergite occurs in Melanthripidae and Merothripidae by the complete loss of the eighth abdominal sternite, although the two pairs of setae that are associated with the eighth sternite in the other two families are retained submarginally on the seventh sternite of all female Aeolothripidae.
The sensoria on antennal segments III and IV are linear in most Aeolothripidae with a slight curve around the apex of the segments. The antennal sensoria of Rhipidothrips are interpreted as having almost lost the linear portion and retained only the distal curved portion, but R. gratiosus retains the linear portion of both sensoria as a series of isolated areas (Hoddle et al., 2008). Species in three genera do not have linear sensoria, these being Cycadothrips in Australia, and Dactuliothrips and Orothrips in western USA.
Bhatti (2006) proposed that the Aeolothripidae should be recognised as a superfamily Aeolothripoidea. Moreover, he proposed removing seven genera from the family, and placing each into its own family. He thus recognised eight families for this group, seven each with a single genus and Aeolothripidae with 16 genera. This essentially phenetic classification was discussed by Mound & Morris (2007).